
Kıtaların Kayması ve Canlı Dağılışına Etkisi
Cansu Çulha[1]
Giriş:
Dünyayı güneş sistemi içerisinde yer alan öbür gezegenlerden farklı tutan en önemli hususlardan biri, kıtaların kayması ya da diğer adıyla levha tektoniği aracılığıyla sürdürülen jeolojik evriminin var olmasıdır (Polat, 2016). Dünyamızdaki bu jeolojik olayların geneli, levha sınırlarında oluşan fiziksel ya da kimyasal olaylardan etkilenme yoluyla meydana gelmektedir. Bu etkileşimler sonucunda yeni dağ sıraları ve adalar oluşurken; Asya ve Hindistan kıtalarının birleşimi gibi kıta birleşimi ya da Afrika ve Güney Amerika kıtalarının ayrımı gibi kıta ayrımı da gerçekleşmektedir. Ayrıca Alaska ve Batı Kanada gibi de bazı adaların kıtalara ek olarak yerleştiği de görülmektedir (Polat, 2016).
Levha sınırlarında oluşan bu olaylar neticesinde, canlıların yer aldığı coğrafik ortam diğerleri ile ilişkisi kesilmekte veya oluşan yeni ilişkiler ile canlılarda göç etme durumu görülmektedir. Yaşanılan coğrafik ilişki kesintileri, göç etmeleri ve ortam değişikliği, türlerin bazılarının yok olmasına ya da yeni türlerin meydana gelmesine sebebiyet vermektedir (Polat, 2016).
Jeolojik Evrim ile Canlı Dağılışı:
Paleozoik sonları ile mezozoik döneminin başlarında var olan Süperkıta Pangea’nın parçalanmaya başladığı süreçte, okyanusal alanları genişlemiş ve Orta ve Güney Atlantik okyanusunun gelişiminin yanı sıra Kuzey Atlantik oluşumu da başlamıştır. Bu süreç içerisinde Kuzey Amerika’da yer alan büyük iç deniz kurumuş ve Grönland’da Amerika kıtasından ayrılmıştır ve Güney yarım kürede Gondwana kıtası parçalanmalara devam etmekte ve Avustralya bu kıtadan ayrılmıştır. Bu değişiklikler gerçekleşirken ormanların geneli, güneyde yer alan kıtalarda; tropikal ve yarı tropikal yağmur ormanları ise ekvator çevresinde yer kaplamıştır. Bu bölgelerin yanı sıra kuzeyde daha soğuk alanlarda çam ormanları yayılış göstermiştir (Sakınç, 2006).
Amerika kıtasının kuzey ve güneyi, evrimleşmeye başlayan marsupial türündeki keseli hayvanlara kuzey bölgelere gitmesi için yol meydana getirmiştir. Bilimsel verilerden elde edilen kanıtlar doğrultusunda fosiller görüşü doğrular niteliktedir. Bu durumda plesental memeliler ise kuzeyde yer alan kıtalarda kemirici ve primatlar ile yayılımını sürdürmüştür. Aynı şekilde fosil kanıtlar doğrultusunda Güney Amerika’da yer alan toynaklıların kuzeyde ortaya çıktıktan sonra güneye karaköprü sayesinde geçiş yaptıkları belirtilmektedir (Sakınç, 2006).
Bu durumlar neticesinde günümüzde uzaklıkları oldukça fazla olan kara parçaları 500 milyon yıl önce birleşik vaziyette bulunduğu anlaşılmaktadır. Büyük yok oluşun ardından ortama adapte olabilen bazı faunalar evrimleşip hayatta kalmaya devam etmişlerdir. Böylelikle 500 milyon yıl önce evrimleşen bu faunalar çeşitlenmeler ile milyonlarca yıllık bir yolculuğa adım atmıştır (Sakınç, 2006).
50 milyon yıl kadar önce ya da Eosen döneminde Kuzey Amerika ile Asya kıtası arasında oluşan Bering Köprüsü sayesinde hayvanlar bu iki kıta arasında dağılış göstererek önemli bir coğrafik olayı oluşturmuştur (Türkeş,2015). Bu olay deniz suyu seviyesinin düşmesi ya da kara parçalarının tektonik nedenlerle yükselmesi sonucunda kıtaların arasında karasal bağlantı meydana gelmiş ve biota alışverişi Bering kara köprüsü ile dağılış göstermiştir (Demirsoy,2002).
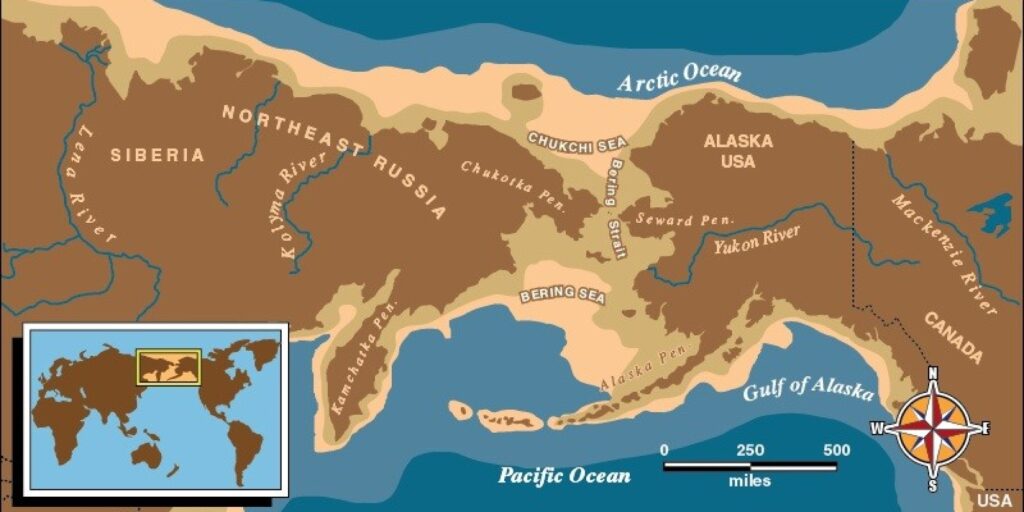 Görsel 1. Beringia’yı gösteren açık kahverengi bir kenarlığa sahip doğu Rus ve Alaska haritası (13).
Görsel 1. Beringia’yı gösteren açık kahverengi bir kenarlığa sahip doğu Rus ve Alaska haritası (13).
Bugün bir kara köprüsü Panama kanalında Kuzey ve Güney Amerika’yı birbirine bağlamaktadır. Ancak Eosen döneminin ortasından (günümüzde 50 milyon yıl) Pliyosen döneminin sonuna kadar (günümüzde 3 milyon yıl), iki kıta tamamen sularla ayrılmıştır. Bu uzun dönem boyunca, büyük memeli grupları her kıtada farklı yönlerde evrimleşmiştir. Pliyosen döneminin sonunda kara köprüsü yeniden kurulduğunda, her iki yönde de bir memeli dalgası akmaya başlamış ve bu dağılma, dünya tarihinde farklı kıtasal faunaların en önemli karışımlarından biri olan “Büyük Amerikan Değişimi” olarak adlandırılmıştır (10).
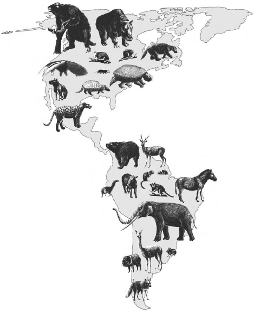
Görsel 2: Büyük Amerikan Değişimi.
En üstte, kıta boyunca kuzeye doğru yürüyen 38 Güney Amerika cinsinin temsilcileri bulunmaktadır. Altta ise, Güney Amerika’ya göç eden 47 Kuzey Amerika cinsinin temsilcileri bulunmaktadır. Kuzey Amerikalı göçmenler, Güney Amerika’ya girdikten sonra hızla çeşitlenmiştir. Kuzey Amerika’ya giden Güney Amerikalı göçmenler çok az çeşitlenmiş veya çoğu yok olmuştur (10).
Karaköprü dönemine girildiğinde izolasyonların ortadan kalkması ile göçler fazlalaşmakta ve çeşitlenmeye sağlanmaktadır. Bu durumda yarı tropikal ormanlar her iki kutupta da yayılım göstermekte olup kuzey enlemlerinde Amerika ve Avrasya’da yağmur ormanları yer almaktadır. Güney enlemlerde tropikal ormanlar Amerika, Afrika ve Avustralya kıtalarında yayılış göstermektedir. Ayrıca geniş ormanların bu denli yaygın olması memelilerin gelişimine katkıda bulunmuştur. Eosen döneminde Memelilerin yayılışlarında en ön safhalarda genellikle kemiriciler yer almaktadır. Fosil kanıtları sayesinde de toynaklı ve keseli memelilerin de yayılışta önem sarf eden bir durumda yer aldığı bilinmektedir (Sakınç, 2006).
Afrika ve Avrasya kıtalarının yaklaşımı sonucunda ise Tetis Okyanusu kapanmaya başlamış ve sonrasında Pratetis Denizi yok olmuştur. Bu durumun yanı sıra Asya ve Avrupa arasında yer alan Turgay Denizi de Eosen döneminin sonlarında kapanmaya başlamıştır. 30 milyon yıl öncesinde denizlerin ortadan kalkması coğrafik engeli aşmış ve Asya’da kalmış olan memelilerin Avrupa’ya dağılımı sağlanmıştır. Diğer bir yandan bu göçü tetikleyen unsur Alp Dağları’nın yükselmesi ile oluşmuş Dinarid-Pelagon-Küçük Asya karaköprüsü olmuştur. Antrakoterıum ve diğer canlılar, Trakya bölgesinden güney kesimlere doğru bu karaköprüsü ile dağılım göstermiştir. Edirne çevresinde bu memeli türünün fosillerine ulaşılarak Avrupa’dan güney kesimlere göç ettiği kanıtlanmış bulunmaktadır (Sakınç, 2006).
Avustralya ve Güney Amerika kıtalarında yaşayan bölgeye mahsus memeliler kıtaların ayrılmasının ardından ortama uyum sağlamak için gelişim göstermişlerdir. Bu canlılardan tavşangiller düzlüklerde hayatını sürdürmeye adapte olmuş ve bitkiler aracılığıyla beslenmelerini sağlamışlardır. Paleosen dönemindeki ormanlarda görülen toynaklı kurt bu değişiklere uyum sağlayamayarak yok olan türler arasında yerini almıştır (Sakınç, 2006). Bu durumda kıtaların kayma teorisi nedeniyle yeryüzünde oluşan coğrafik değişiklikler ile organizmalar popülasyonlarının ayrılması ve yok olması meydana gelmiştir. Kıtaların bu denli ayrılması neticesinde bazı bilim insanlarının tabiriyle kendi kargolarını da yanında götürmüşlerdir[2] (Avcı, 2011).
Miyosen döneminde Afrika kıtası Avrupa kıtasına yaklaşımını sürdürürken Tetis okyanusu da tükenme sürecine girmiş bulunmaktadır. Tetis Okyanusu’nun, Arabistan Yarımadasının Küçük Asya bölgesinin güneydoğusu ile çarpışması sonucunda Hint Okyanusu ile bağlantısını koparılmaktadır (Tassy, 1990). Bu kara parçaları arasındaki Tetis Okyanusunun ortadan kalkması ile Doğu Afrika vadilerinde yaşamını sürdüren pek çok memeliler Anadolu’nun içlerine ilerleyerek önemli yaşam alanları meydana getirmişlerdir. Böylece Anadolu’da 23 milyon yıldan beri göçler ile gerçek memeliler oluşuma adım atmıştır. Bu göç zamanında zürafa, gergedan, sırtlan, hortumlular, böcekçiller, geyikler, kemirgenler, suaygırıları ve insansıların bu bölgeye olan hareketinin kanıtları araştırmaları halen devam eden fosiller ile sürdürülmektedir (Agusti ve Anton, 2002).
Miyosen zamanında okyanusların yarattığı coğrafik engeller karaköprüsü ile kimi zaman aşılsa da bu özelliğini yitirdiği ve yeniden engel durumuna geldiği bilinmektedir. Bu kesiklik memelilerin hareketini de etkilemekte ve istikrarlı duruma gelmesi halinde kıtaların arasında yapılan geçişlerde karmaşalara yol açmaktadır. 5-2 milyon yıl kadar geriye gidildiğinde dünya şu andaki durumuna benzer hale gelmiştir. Bu durumda Asya ile Kuzey Amerika arasında oluşan Bering Boğazı ve Amerika kıtaları arasında yer alan Panama Boğazı dolaylarında oluşan karaköprüler memelilerin göçlerini hızlandırmış bulunmaktadır (Sakınç, 2006).
Yaşamın halen belirgin olduğu kıtalar haricinde Antarktika kıtası, diğer kıtalardan kopuşunun ardından güney kutbuna doğru ilerlemesi biyoçeşitliliği açısından oldukça önem sarf etmektedir. Antarktika’da elde edilen fosillerle bir dönem bu kıtada da bitki ve hayvanların yaşamı için bir ortamın bulunduğu anlaşılmıştır. Geçmiş dönemlerde biyoçeşitliliğinin zengin olduğu düşünülen bu kıtada da canlı türlerinin tamamı buzullarla kaplandığı için yok olmuştur (12).
Bunlara ilave olarak; bilimsel araştırmalar ışığında tarihte en az beş adet büyük ve daha çok sayıda küçük canlı türlerinin yok oluşla karşılaştığını belirtilmektedir. Dönemsel olarak bakıldığında bunlar; Geç Ordovisiyen, Geç Devoniyen, Geç Permiyen, Geç Triyas ve Geç Kretase jeolojik zamanlarında olduğu görülmektedir (Polat, 2016). Canlı türlerinin bahsedilen şekilde yok oluşlarına, geç Neoproterozoik ve erken Paleozoik zamanlarında varlığını sürdüren Iapetus Okyanusu’nun kıyı kesimlerin derinliği az olan sularında yaklaşık 600 milyon yıllarında yaşamakta olan omurgasız canlılardan Ediakarian hayvan topluluğu örnek gösterilebilmektedir. Ediakarian hayvan topluluğunda kurt, süngerler, denizanası, ilkel eklembacaklılar yer almaktadır. Bu faunanın fosillerine Avustralya’nın güneyinde yer alan Ediakarian Tepeleri üzerinde bulunan kumtaşlarında karşılaşılmıştır. Bunun yanı sıra Ediakarian hayvan topluluğuna ait fosillerin Rusya, Kanada, Çin ve Greenland’de de bulunduğu belirtilmektedir (Sakınç, 2006).
Araştırmalar dahilinde canlıların Kuzey Pangea bozulması sonucunda gerçekleşen yok oluşlarının temelinde üç çevresel nedenin yattığı düşünülmektedir. Birinci neden olarak derin su bölgelerinin kalsit oranlarının derinlere inildikçe artan oksijen yoğunlaşmasıyla yaşanan lisoklin sığlaşması gösterilmektedir. Lisoklin alanlarındaki daralma, kalsitin okyanusun içerisinde zorlu koşullarda çözüme ulaşmasına neden olmaktadır. Bununla beraber dallı bacaklı ve mercan türleri hayatına devam edebilmek için çözünmüş derecede kalsit ihtiyacı duyan karbonat üreticilerin ortadan kalkmasına neden olmuştur. İkinci bir neden Pangea’nın ayrılması sonucunda oluştuğu varsayılan Sibirya volkanlarının lav püskürtmesi olarak görülmektedir. Alandaki volkanik püskürtmeler zehirli metallerin etrafa yayılımıyla kısıtlamalara neden olmuştur. Atmosfere salınan aşırı karbondioksit ile lisoklin alanlarının daraldığı bu nedeni oluşturmaktadır. Son olarak yok oluşta etmen olan üçüncü neden kuzey Pangea başlangıçtaki anoksik ortamı kaynak olarak gösterilmiştir. Fazla yüklenen metal nedeniyle anoksik okyanusların asitleriyle dipteki ihtiyaç duymakta olan türler ortadan kalkmıştır (Bond ve Grasby, 2017).
Ayrıca kıtaların kayması, vikaryans biyocoğrafyası olarak adlandırılan yerkürede aralarında oldukça yüksek mesafeler olan türlerin ve bioatanın pasif durumda taşınışına yol açmıştır. Bu duruma göre türlerdeki dağılımların arasında oluşan parçalanmalar, dağ oluşumuna benzer biçimde gelişen edilgen ayrılmaların neticesini oluşturur. Bu model bir takson veya türün coğrafik olarak dağılışının, yok oluş ve ayrılma durumunda uyum içerisinde göstereceğini varsaymaktadır. Örnek olarak; Doğu Yarım Kürede yer alan Akdeniz Havzasındaki doğu çınarı, Batı Yarım Kürede yer alan kuzey Amerika’da dağılışının batı çınarı türü verilmektedir. Farklı kıtalar içerisinde bulunan grupların vikaryansın bir sonucu olarak farklılaştığı belirtilmektedir. Bu duruma örnek olarak yukarıda bahsedilen karaköprüler de verilmektedir (Avcı, 2011).
Levha tektoniği kuramı ve derin deniz araştırmaları ışığında elde edilen bilimsel verilerle fosillerin önemli bir kanıt teşkil ettiği görülmektedir. Güney Afrika kıtasının batısında ve Güney Amerika kıtasının doğusunda Üst Permiyen Yaşlı Mesozor adını almış su sürüngeninin fosil lokaliteleri sayesinde yaklaşık 250 milyon yıl öncesinde Amerika ve Afrika kıtasının birleşik olduğunu ortaya koymaktadır. Fosiller üzerinden elde edilen verilere bir diğer örnek eğrelti otu bitkisinin 300 milyon yıl kadar öncesinde, Prekambriyen’in sonlarında Antarktika, Avustralya, Afrika, Güney Amerika, Hindistan kıtaları ile Arabistan ve Madagaskar’ın birleşimiyle ortaya çıkan Gondwana kıtasını kaplamaktadır. Günümüzde ise uzaklıkları oldukça fazla olan bu kıtalar ve bölgeler üzerinde fosillerine rastlanmaktadır (Sakınç, 2006).
Bazı kanıtlar, hayvanların mevcut dağılımının kıtasal sürüklenme teorisi temelinde açıklamaktadır. Kuzey Yarım Kürede (Nearktik ve Palearktik bölge) memelilerin dağılımının, güney bölgelerdeki (Afrika, Güney Amerika ve Avustralya) memelilerin dağılımından daha fazla benzerlik gösterdiği bilinmektedir. Bu durum kuzey bölgesinin hayvanlarının son zamanlarda Bering Boğazı’ndaki kara bağlantıları nedeniyle yayıldığını belirtmektedir. Güney Yarım Kürenin memelileri ise, kıtaların parçalanması sırasında ortaya çıkmış ve kıtaların ayrılmasından sonra evrim geçirmiştir. Memeliler izole edildikten sonra farklı şekillerde evrimleşebilmekte ve Gondwana ilk bölünmeye başladığından, farklı kıtalardaki memelilerin ayrı ayrı evrimleştiği ve yayıldığı düşünülmektedir. Bu nedenle, lamalar, alpakalar, tembeller, armadillolar gibi Güney Amerika memelileri, Güney Yarım Kürenin diğer kıtalarında bulunmamaktadır. Etiyopya bölgesinin zürafa, zebra, su aygırı, şempanze ve gorilleri ile Avustralya bölgesinin yumurtlayan memelileri ve bazı keseli türleri bu bölgeler dışında hiçbir yerde bulunmamaktadır. Gondwana’daki memelilerin dağılımı farklı olsa da bunun aksine, bazı balıkların, amfibilerin ve sürüngenlerin dağılımı kıtaların kayması teorisi ile ilgili olarak çok benzerlik göstermektedir (11).
Akciğerli balıkların dağılımı da kıtaların kayma teorisini desteklemektedir. Orta Devoniyen’de ortaya çıkan dipnoanlar, şimdi üç cins Protopterus, Lepidosiren ve Neoceratodus tarafından temsil edilmektedir ve sırasıyla Afrika, Güney Amerika ve Avustralya’da bulunmaktadır. Bu dağılım ile, Orta Devoniyen ve Afrika, Güney Amerika ve Avustralya’daki Gondwana topraklarının o dönemde temas halinde olduğunu ve bu kıtaların ayrılmasıyla şu anda dipnoanların üç ayrı cinsle temsil edildiğini göstermektedir. Amfibiler arasında, üyelerin dağılımı, Kıtaların Kaymasını rastgele desteklememektedir. Sadece Afrika ve Güney Amerika arasındaki pipidlerin dağılımı ve Nearctic ve Avrasya’nın doğu kesimlerindeki semenderler kıtaların kaymasını bir dereceye kadar desteklemektedir. Bunların yanı sıra sürüngenler arasında da bazı üyelerin dağılımı kıtaların kaymasını desteklemektedir. İlkel kaplumbağaların bazı üyeleri Permiyen döneminde ortaya çıkmış ve hala Güney Yarım Kürede sınırlı durumda kalmışlardır. Timsahlar ve gavyal gibi timsahlar, şimdi Orta Amerika, Afrika ve Kuzey Avustralya bölgesinin bazı alanlarıyla sınırlı olan Triyas döneminde ortaya çıkmıştır (12).
Anadolu yarımadasına bakıldığında ise geçmiş yıllarda gerçekleşen göçler sayesinde dünyanın en güneyi ile en kuzeyinde yer alan bitkilerin dağılımı görülmüştür. Bu duruma örnek Afrika kıtası ve Asya kıtasının güneybatı kısmında bulunan karakulak ve çizgili sırtlanların dağılımlarının sınırlarından en kuzeyi Anadolu Yarımadası oluşturmaktadır. Benzer biçimde bozayı ve sarıçamlarında dağılış alanları Avrupa ve Sibirya olmakla beraber sınırının en güneyini Anadolu oluşturmaktadır. Anadolu bu sebeple tür açısından zengin ve de uç yayılış içeriği açısından genetik çeşitliliği önemli bir konumdadır. Paleoiklimsel değişikliklerin ve Anadolu topoğrafyasının doğurduğu sonuçlarla beraber bu topraklarda kurbağa ve sürüngenlerin çeşitlenmiş olması önemli rol oynamaktadır. (Tavşanoğlu, 2016)
Jeolojik değişiklikler yaklaşık 200 milyon yıllık bir zaman diliminde meydana gelmesine rağmen, bazı organizmalar evrimsel değişime dair çok az kanıt göstermektedir. Örneğin, yan boyunlu kaplumbağalar (Pleurodira alt takımı) Jura döneminden beri çok az değişmiştir. Bilim insanları, kıtasal parçalanma yaşanmadan önce Gondwana’ya dağıldıklarını varsaymaktadırlar. Bugün bu türdeki kaplumbağalar Güney Amerika, Afrika, Madagaskar, Avustralya ve Hint Okyanusu adalarının bazı bölgelerinde bulunmakta, ancak dünyanın başka hiçbir yerinde bulunmamaktadırlar (Gibbons, 2020). Wegener’in yaptığı birçok çalışma neticesinde de bahsedilen bu düşünceler doğrulanmıştır. Bilhassa solucan ve yengeçlerin evrimini az yaşamış olarak şu anki döneme kadar gelmiş olması kıtaların kaydığının önemli bir kanıtıdır.
Kara kütlelerin bir araya gelmesi de biyoçeşitlilik için büyük önem sarf etmektedir. Örneğin Afrika kıtası ile Hindistan levhası birbirinden ayrılarak Avrasya ile birleştiği sırada Afrika ile Hindistan arasındaki canlı bağları kesinti yaşamış, Hindistan ile Avrasya’da yaşayan canlılar arasında ise etkileşimler yaşanmıştır. Kıtaların kayması ile biyoçeşitliliğe etki eden bir diğer durum ise deniz altındaki tabakaların su yüzeyine çıkarak kara yaşamına elverişli hale gelmesidir. Örnek olarak, Avrasya ile Afrika, Arabistan ile Hindistan levhalarının arasında bulunan Tetis Denizi’nin tabanındaki tortul tabakaların su yüzeyine çıkması verilebilir. Bunun sonucunda Tetis Denizi’nin ekosistemi ortadan kalkmış ve su ekosistemi kara ekosistemine dönüşmüştür. Tam tersi biçimde kara alanlarını sular altında kaldığı yerlerde mevcuttur. Bu durumda, Türkiye ile Yunanistan’ın arasında bulunan Egeid kara parçasının çökmesi ile deniz meydana gelmiştir ve sonucunda kara ekosistemi yok olup su ekosistemine dönüşmüştür (12).
KAYNAKÇA:
- Agusti, J., Anton, M., Mammoths, Sabertooths, and Hominids: 65 Million Years of Mammalian Evolution in Europe, Columbia University Press, 2002.
- Avcı, M. Moleküler Biyocoğrafya: Gelişimi, Kapsamı, Paleobiyocoğrafya ve Biyolojik Çeşitlilik Açısından Bir Değerlendirme, Fiziki Coğrafya Araştırmaları: Sistematik ve Bölgesel, Türk Coğrafya Kurumu, s.241-266, 2011.
- Bond D., Grasby S., On the causes of mass extinctions, Palaeogeography, Palaeoclimatology, Palaeoecology, Sayı 478, s. 3-29, 2017.
- Demirsoy, A., Genel Zoocoğrafya ve Türkiye Zoocoğrafyası “Hayvan Coğrafyası”, Meteksan Ankara, 2002.
- Polat, A., Yerküre üzerindeki yaşamın kökenine ve evrimine jeolojik bir bakış açısı. Popüler Yerbilim Dergisi, 2016.
- Sakınç, M., Jeoloji ve Biyolojik Evrim iç içe: Yerin Evrimi. Bilim ve Gelecek Dergisi, Sayı 26, s. 8-33, 2006.
- Tassy, P., The “proboscidean datum event”: how many proboscideans and how many events? in: Lindsay e.H., Fahlbusch v., Mein P. (eds.). European Neogene Mammal Chronology, Plenum Press, New York, s.237-252, 1990.
- Tavşanoğlu, Ç., Anadolu’nun yüksek biyoçeşitliliği: Evrim bunun neresinde? N. E. Iraz Akış (Dü.) içinde, Evrimin Işığında, Yazılama Yayınevi, s. 207-225, 2016.
- Türkeş, M., Biyocoğrafya: Bir Paleocoğrafya ve Ekoloji Yaklaşımı, Kriter yayınevi, Ankara, 2015.
Web Siteleri:
- https://biocyclopedia.com/index/general_zoology/continental_drift_theory.php
- https://www.biologydiscussion.com/animals-2/phylum-chordata/continental-drift-theory-evidences-and-explanation-biology/40477
- https://www.cografyaci.gen.tr/biyocesitliligi-etkileyen-paleocografya/
- https://www.nps.gov/bela/learn/historyculture/the-bering-land-bridge-theory.htm
[1] Gazi Üniversitesi Eğitim Bilimleri Enstitüsü Coğrafya Eğitimi Yüksek Lisans Öğrencisi.
İTÜ Avrasya Yer Bilimleri Enstitüsünde Yürütülen ” Genadaptif: Türler Arası Gen Geçişlerinin Türlerin Adaptif Potansiyeli Üzerine Etkilerinin Ekolojik ve Genomik Olarak Araştırılması” adlı projede yüksek lisans düzeyinde araştırmacı.
[2] Bahsedilen kargo için yaşamını devam ettirmekte olan canlılar ve fosil kanıtlar örnek olarak verilmektedir.